

Do Wetlands Filter Acid Mine Pollutants
Introduction
Pollution from abandoned mine sites is a major ecology trouble that has deleterious consequences for aquatic and terrestrial ecosystems (Bridge, 2004). Freshwater resource are negatively impacted by mining contagion (Johnson and Hallberg, 2005), reducing the value of h2o for agriculture, recreation, or manufacture, and rendering it unsafe for human consumption (Tripole et al., 2006). Release of metals from exposed rock and tailings plus uncontrolled drainage of metal-contaminated h2o is a common challenge from disused mines. For many mines, including coal mines, the drainage is also highly acidic. This acrid mine drainage (AMD) is generated when metallic sulfide minerals, mainly pyrite, are oxidized in the presence of water and accelerated by the action of chemolithotrophic bacteria. This generates acerbity and mobilizes large concentrations of iron (Fe), and other toxic trace metals and metalloids, depending on the composition of the exposed rock (Robb, 1994; Stonemason, 2002). AMD pollution from abandoned mines worldwide therefore is a major trouble that needs to exist mitigated and managed (Hedin et al., 1994; Mayes et al., 2009b).
The label of microorganisms in such polluted environments is important in society to empathise the impacts of AMD on microbial ecology and development, to place microorganisms that may accept bioremediation properties, and to understand the mechanisms of microbial AMD tolerance and remediation (Méndez-García et al., 2015; Chen et al., 2016; Huang et al., 2016). The ability to identify bacterial taxa and quantify bacterial abundance and diversity in AMD environments has been significantly advanced through the employ of adjacent-generation sequencing and metagenomics tools (Amaral-Zettler et al., 2011; Kuang et al., 2013; Brantner and Senko, 2014; Liu et al., 2014). In particular, this has allowed bacterial community structure and water quality status to be correlated with functional characteristics (Chen et al., 2015; Hua et al., 2015; Kuang et al., 2016). These analyses revealed that while AMD environments accept significantly reduced species richness and diverseness, they exhibit a high abundance of acidophilic taxa. Moreover, bacterial diverseness betwixt dissimilar AMD environments can exist predicted in part by physicochemical characteristics, particularly pH (Kuang et al., 2013; Liu et al., 2014). These studies also demonstrate that AMD tolerant bacteria use multiple metabolic processes to survive these environments, such every bit high expression of genes encoding fe and sulfur oxidation enzymes, multiple modes of carbon and nitrogen metabolism, and loftier abundance of metal transporter genes to withstand metal stress. Furthermore, in that location is prove that transcriptional profiles of bacterial communities alter in response to dissimilar AMD water quality characteristics (Chen et al., 2015; Kuang et al., 2016).
Microbial community structures have been examined in mine tailings, AMD water and sediment, and biofilms from mine sites (Huang et al., 2016). However, very few previous AMD studies have used large-scale sequencing approaches to examine leaner associated with wetland found rhizosphere sediments (Diaby et al., 2015), and to our knowledge, none have characterized AMD-exposed natural wetlands in direct comparison to river sediment without wetland plants. Wetlands tin prove substantial resilience to highly metal-rich and acidic waters over very long periods of time; for example, natural wetlands that accept been studied in Republic of ireland, United Kingdom, and United States take been plant to tolerate very high acidity (sometimes pH < 3), loftier dissolved metal concentrations (such as >200 mg L-1 of Fe) over many decades, indicating long-term adaptation to AMD stress (Beining and Otte, 1996; August et al., 2002; Dean et al., 2013). Indeed natural and constructed wetlands are oftentimes considered every bit a passive, low-maintenance approach to mine drainage remediation by slowing and reducing drainage run-off, enhancing bulk uptake of metals into biomass, and past providing organic carbon to maintain rhizosphere bacteria to bulldoze redox reactions including sulfate reduction (Sheoran and Sheoran, 2006; Mayes et al., 2009a; Dean et al., 2013). These reactions volition in turn reduce concentrations of dissolved metals through the germination of metal precipitates and sedimentation, and promote alkalization (Hallberg and Johnson, 2005). Nevertheless, it is argued that wetlands are complex systems that are not an appropriate solution for AMD remediation if not properly managed or understood (Johnson and Hallberg, 2002). A more cardinal understanding of the microbiological processes that are taking place within a wetland is therefore essential in social club to better elucidate the mechanisms for improved remediation. This study aimed to enhance this understanding by quantifying microbial customs dynamics within an AMD-impacted natural wetland, and to specifically address the hypothesis that differences in microbial customs composition and metabolic activities of these communities can account for the remediation observed in the wetland.
The United Kingdom has a long history of metallic mining leading to significant pollution problems as a result of AMD (Mayes et al., 2010). The Parys Mountain (Mynydd Parys) copper mine in Anglesey, Northward Wales, ceased operation in 1911 but accounts for some of the highest releases of Atomic number 26, Cu, and Zn nationally (Mayes et al., 2010). Significant volumes of AMD enter the small northern Afon Goch river via a mine crawlway (Effigy 1). Following changes in drainage routes from the mine, this river has experienced substantially greater pollution in the last 15 years such that information technology is a recently impacted ecosystem that has non fully adapted to AMD with no abundance of wetland plants (Coupland and Johnson, 2004). AMD also enters a second river, the southern Afon Goch, which has experienced AMD pollution for over a century. While heavily polluted by dissolved metals and acidity at its source, in that location have been improvements to downstream water quality in this river due partly to remediation past a large natural wetland (Boult et al., 1994; Batty et al., 2006; Dean et al., 2013). This river system provides an ideal written report site to quantify and contrast the microbial community limerick in river sediment of a recently impacted ecosystem without wetland vegetation compared to a long-term impacted ecosystem with a substantial natural wetland, which is expected to take adjusted to AMD pollution. In addition, past comparing the AMD affected wetland with a nearby unpolluted wetland site, we are able to examine changes in wetland microbial community structure on the basis of AMD exposure. This will let us to test our hypothesis and examine whether in that location are differences in predicted metabolic activities derived from distinct bacterial communities found in wetland and river sediment sites.
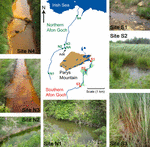
Figure one. Sample sites within the Parys Mountain river catchment in Anglesey, Wales, United Kingdom. Sites on the southern Afon Goch river are marked S1–S3, with the wetland areas shaded in green. Sites on the northern Afon Goch river are marked N1–N4. Representative photographs of the samples sites along each river taken during times of sampling are shown. NA is the location of the adit that discharges AMD to the river.
Materials and Methods
Field Site Locations and Sampling
H2o and sediment samples were collected from five sites forth the northern Afon Goch river and iii sites within the wetland through which the southern Afon Goch river flows (Figure 1). Samples were likewise taken at an unpolluted wetland (UW) site at Cefni Reservoir that is 13 km s of Parys Mount. Exact locations are provided in Supplementary Tabular array S1. The northern Afon Goch rises to the west side of Parys Mount and runs approximately v.5 km in length with the Dyffryn Adda adit from the Parys Mountain mine joining at approximately 2.5 km along the river. The river then runs n through the town of Amlwch and enters the Irish gaelic Ocean through an industrial site next to the port. The river course is largely canalized. Five sites were sampled (Figure 1) two upstream of the crawlway (site N1 and N2), the Dyffryn Adda adit itself (site NA), and two sites downstream of the crawlway (N3 and N4). Sampling was carried out on four (at site N1) or vi (sites N2–N4 and NA) occasions on June 2010 (all sites), July 2010 (all sites), August 2013 (all sites), October 2013 (non site N1), March 2014 (all sites), and October 2014 (not site N1).
The southern Afon Goch is xi km in length and runs south of Parys Mountain (Figure ane). Approximately 500 m beneath the mine source at the Mona crawlway, the river flows through a natural wetland of ∼0.i km2. The river and then runs east and enters the Irish Sea at Dulas Bay. Three sites were sampled within the wetland (sites S1–S3). Site S1 is an entry point for AMD runoff at the showtime of the wetland, while sites S2 and S3 are in the middle and at the end of the wetland, respectively (Figure ane). Sampling was carried out on four (site S2) or 7 (sites S1 and S3) occasions on June 2010 (all sites), July 2010 (all sites), Nov 2011 (all sites), August 2013 (all sites), October 2013 (non site S2), March 2014 (all sites), and October 2014 (not site S2).
Triplicate sediment and water samples for analysis of pH, electrical conductivity (as a measure out of conductive ion concentrations), and dissolved, sediment, and particulate metals were collected at each site on each sampling occasion. A YSI 556 probe (Xylem Analytics, Letchworth, United Kingdom) was used to monitor water pH, conductivity, and dissolved oxygen. Samples for metallic analysis (dissolved, sediment, and particulate) were nerveless at each site and processed within five h following drove. For dissolved metals, a known book of river water (typically 100–200 mL) was filtered through a 0.45-μm cellulose acetate filter and the filtrate was stored in an acid-washed polypropylene canteen and was acidified to ii% with ultra-pure nitric acrid. The pre-weighed filters were retained for analysis of metal particulates. Sediment samples were collected to approximately 1 cm depth using a plastic scoop and sealed in a plastic bag until analysis. On return to the laboratory, the filters containing suspended particulates were dried (60°C for 48 h), and the sediment samples were dried (threescore°C for 48 h) and and so passed through a 250-μm filter. The particulate filter papers and 100 mg of sediment samples were digested in 67% ultra-pure nitric acrid for 24 h at 100°C. Digests were and so diluted to 2% nitric acid in deionized Milli-Q h2o (Millipore). Samples were refrigerated before metallic concentrations in water samples and digests were analyzed by inductively coupled plasma atomic emission spectroscopy (ICP-AES) using a Perkin-Elmer Optima 5300 for Al, Equally, Cd, Cu, Iron, Mn, Atomic number 82, and Zn. The spectroscope was calibrated using an internal standard, which was a matrix matched series dilution of Specpure multi element plasma standard solution 4 (Alfa Aesar).
For microbial customs assay, 3 sediment samples were nerveless from each site in March 2014 for DNA extraction. The southern Afon Goch wetland and Cefni Reservoir wetland are both dominated by Juncus sp. and samples were taken from sediment surrounding Juncus sp. roots from sites S1, S2, S3, and UW. In addition, a non-vegetated sediment sample was taken from site S1 (S1R), and a sediment sample surrounding cotton fiber grass (Eriophorum angustifolium) roots was also taken from site S2 (S2C). Samples were taken in sterile l-mL Falcon Tubes.
Deoxyribonucleic acid Extraction and 16S rRNA V3–V4 Cistron Amplicon Sequencing
DNA was extracted from 100 mg sediment samples (iii independent samples per site) using a Powersoil Deoxyribonucleic acid isolation kit (MoBio Laboratories, Carlsbad, CA, United States) and quantified using a Nano-drop 3300 (Thermo-Scientific, Waltham, MA, United States). DNA from each site was pooled and then amplicons for Illumina MiSeq sequencing were generated from PCR reactions using primers Bakt_341F (S-D-Bact-0341-b-South-17) and Bakt_805R (Due south-D-Bact-0785-a-A-21) as previously described and validated (Herlemann et al., 2011; Klindworth et al., 2013), modified with Illumina overhang adaptors (forward overhang: 5'-TCG TCG GCA GCG TCA GAT GTG TAT AAG AGA CAG; reverse overhang: v'-GTC TCG TGG GCT CGG AGA TGT GTA TAA GAG ACA Thou), to amplify the V3–V4 region of the 16S rRNA gene. PCR reactions were performed using KAPA HiFi HotStart Mix (KAPA, Woburn, MA, United States) and PCR distension conditions of 95°C for 3 min, and then 25 cycles of 95°C for 30 s, 55°C for 30 south, 72°C for 30 s, and then 72°C for v min. Following purification using AMPure XP beads (Beckman Coulter, High Wycombe, United Kingdom), index PCR and addition of Nextera sequence adapters was performed using a Nextera XT Index kit (Illumina Inc., San Diego, CA, The states) according to manufacturer's instructions. Amplicons were sequenced using an Illumina MiSeq at the Genomic Technologies Facility, University of Manchester, and to a depth of over 100,000 sequences for each sample, and up to one,473,986 for sample N4 (encounter Table ane). Sequence data were deposited in the European Nucleotide Archive (ENA), study accession number: PRJEB23187, sample accession numbers: ERS1983433 (site UW), ERS1983434 (site S1), ERS1983435 (site S2), ERS1983436 (site S3), ERS1983438 (site S2C), ERS1983439 (site N1), ERS1983440 (site N2), ERS1983441 (site NA), ERS1983442 (site N3), ERS1983443 (site N4), and ERS1983444 (site S1R).

TABLE 1. Summary of 16S rRNA gene amplicon sequence analysis and bacterial community diversity parameters for each sample site.
Statistical Analysis of Environmental Information
Principal component analysis (PCA) of environmental data was performed using the R vegan parcel v.ii.4.2. The environmental information (except pH) was log-transformed prior to PCA. The PCA comprised a matrix of 46 datasets (replicates) from the six samples with 24 variables used. Statistical comparison of ecology information was performed using one-mode ANOVA (p < 0.05) and Tukey's multiple comparison mail service hoc exam using GraphPad Prism. Hierarchical clustering (HC) of environmental information was performed in R and statistical comparison of nodes performed by a Similarity Profile (SIMPROF) test (Clarke et al., 2008).
Sequence Data Analysis
Raw de-multiplexed sequence reads of the 16S rRNA V3–V4 region were trimmed and paired and further filtered using QIIME v.1.ix.0 (Caporaso et al., 2010). Chimeric sequences were identified and removed past UCHIME 5.4.2 (Edgar et al., 2011) earlier operational taxonomic units (OTUs) picking. OTUs were de novo picked at 97% similarity using UCLUST (Edgar, 2010). Taxonomic classification of the representative sequences was performed using the Greengenes v.xiii.viii 97% OTU dataset and using the Naive Bayes car learning classifier, which was trained using the amplified V3–V4 region sequence reads (Werner et al., 2011). In order to evaluate the microbial blastoff diversity in each sample, Shannon–Weiner diversity alphabetize (H), Pielou'due south evenness index (H/Hmax ), and Chao1 species richness estimate values were calculated using QIIME. Before the calculations, samples were resampled to an fifty-fifty depth of 310,833 sequences per sample, the sequence depth size of the smallest sample (N2). Rarefaction curves were also generated by QIIME.
The assigned taxa dataset was statistically analyzed by HC and by non-metric multidimensional scaling (NMDS) both performed using R vegan packet v.2.four.ii. Taxa relative abundance was square-root-transformed and a distance matrix based on Bray–Curtis dissimilarity was obtained for HC. Consummate linkage was used as an agglomerative clustering method and SIMPROF was used for identifying 18-carat groups betwixt samples in HC. Samples were classified as "unpolluted" and "polluted" groups according to the ecology information. Analysis of similarity (ANOSIM; Clarke, 1993) was used to evaluate taxa aggregation departure between those groups and similarity percentage (SIMPER) analysis (Clarke, 1993) was used to identity taxa discriminating these groups and their contribution. BIO-ENV (Clarke and Ainsworth, 1993) was used to obtain the all-time subsets of environmental variables explaining taxon aggregation.
Metabolic structure was predicted from the 16S rRNA gene amplicon library of each sample site using the PAPRICA v.0.4.0 metabolic inference pipeline (Bowman and Ducklow, 2015). For this analysis, libraries were subsampled to 310,833 reads, the size of the smallest library. Abundance of enzymes and metabolic pathways were normalized according to the estimated number of 16S rRNA gene amplicon copies for each sample. For generating relative log2 fold-change abundance values, normalized enzyme abundance values were converted to logarithmic calibration (base 2) after adding a value of 10 to allow consideration of zero values. HC of normalized metabolic pathway abundances was performed using the Euclidean altitude method and heatmaps were created using R vegan package v.2.4.2.
Results and Discussion
Environmental Characterization
The southern and northern Afon Goch rivers are both exposed to AMD but have very different environmental characteristics, due in part to the presence of the natural wetland on the southern river, which is dominated by Juncus sp. and some E. angustifolium (cottongrass) and Phragmites sp. The northern Afon Goch has neutral pH and low conductivity water at sites N1 and N2 upstream of the adit entering the northern river, while highly acidic (pH 2.3–two.5), loftier conductivity water was downstream, although this was slightly diluted at site N4 (Figure two). An increment of pH from 2.four to 5.7 and a decrease of conductivity from i.1 to 0.iii mS were observed forth the wetland from site S1 to S3, indicative of the remediation process previously described (Dean et al., 2013). Dissolved metal concentrations inside the h2o across the sites mirror the conductivity profile, and near dissolved metals show a like profile across the sites with highest concentrations at site NA and N3 (Figure 2C). Many particulate metals including Al, Cu, Fe, Pb, and Zn bear witness loftier concentrations inside the centre of the wetland at site S2 (Supplementary Figure S1). This is probable due to the increase in pH causing increased atmospheric precipitation of metals at site S2, in contrast to the college concentration of dissolved metals, and lower particulate metals, within the acidic conditions at site S1. Overall, metallic concentrations inside the sediment are high in the acidic sites, particularly at N3 and N4 (Figure 2E) although sediment metal profiles vary depending on the element (Supplementary Figure S1). One unexpected pattern was the loftier concentration of some metals within sediment (particularly Mn, Pb, and Zn) at site N1 despite this otherwise unpolluted site being located upstream of the mine crawlway. This site tin feel metal rich run-off from the mine site during high rainfall and is a slow-flowing, neutral pH section of the river, which volition lead to greater sedimentation of metals.
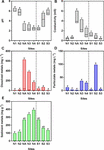
Figure two. Water chemistry [(A) pH and (B) conductivity] and full metal concentration in water [(C) dissolved and (D) particulate] and (East) sediment samples. Information are pooled from triplicate analyses taken on iv to seven sampling occasions (June 2010, July 2010, November 2011, August 2013, Oct 2013, March 2014, and October 2014). Boxes prove the minimum and maximum values and the line within the boxes shows the median values. Boxes that practise not share lowercase letters are significantly different (p < 0.05) as determined by one-way ANOVA. Data for individual metals (Al, Equally, Cd, Cu, Fe, Mn, Pb, and Zn) are shown in Supplementary Effigy S1.
Principal component analysis indicates that the loftier concentrations of dissolved metals are primal variables that correlate with the clustering of sites NA, N3, and N4, and to a lesser extent with site S1 (Figure 3). In dissimilarity, the less polluted sites N1, N2, and S3 correlate with high water pH (pH 5.1–6.4), high concentration of Mn in sediments, and depression concentration of As and Fe in sediments. S2 amassed separately from the other sites (Figure 3A) and correlates with particulate metals in the h2o column (Figure 3B). It was previously observed that the wetland remediation process is mediated in part past atmospheric precipitation of metals in the heart of the wetland (Dean et al., 2013), as also seen here by the particulate metal profile (Figure 2D). Natural wetlands such as this one tin function as long-term metal sinks by reducing period charge per unit, reducing acidity, and promoting microbial activities such every bit sulfate reduction (Beining and Otte, 1996; Webb et al., 1998; Dean et al., 2013). This activeness gives ascent to a subsequent immobilization of metals in the lower reaches of the wetland. In contrast, no significant change in particulate metals is observed downstream of the northern Afon Goch adit, which does not pass through any vegetation.
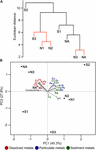
Figure 3. Discrimination of sites on the basis of physicochemical parameters. Hierarchical cluster analysis (A) and PCA ordination plot (B) illustrating the discrimination between sample sites co-ordinate to environmental properties and their correlation with each environmental factor analyzed. In (A), sites with non-significant clustering (p < 0.05) every bit determined past SIMPROF are indicated with red lines.
Microbial Community Construction
DNA from sediment samples at all sites, equally well as from an unpolluted wetland nearby (site UW; Supplementary Tabular array S2) were used for 16S rRNA factor amplicon sequencing. Following processing 367,239–1,473,986 sequences were derived from each site, yielding 3678–21,438 total OTUs (Table 1). All samples showed a consistent level of saturation of OTU richness (Supplementary Figure S2). The crawlway (site NA) had the everyman OTU number whereas the equally highly polluted site N4 had the highest number of total OTUs. Likewise, both the Shannon–Weiner diversity alphabetize and Chao1 species richness gauge indicated that site NA had depression bacterial variety, while the Pielou's alphabetize score indicated that this site had low community evenness. In contrast, unpolluted sites UW, N1, N2, and S3 were the most diverse. In particular, the Shannon–Weiner diversity score indicated that site S3 diversity was essentially improved compared to sites S1 and S2 at the lower reaches of the wetland. Yet the impacted wetland sites S1 and S2 maintained diversity (with scores of 5.73 and 6.56, respectively) compared to the AMD impacted river sites NA, N3, and N4 (with scores of 4.29, 3.36, and v.63, respectively). This pattern was likewise seen with the Chao1 species richness and the evenness scores and demonstrates that the outcome of pollution on species composition is greater in the river sediments than within the wetland. The reduced bacterial diversity at polluted river sites NA, N3, and N4 was besides validated on the basis of OTU taxonomic assignments (Supplementary Data Sheet 1) and visualized in Effigy iv. Excluding the unassigned group, the unpolluted sites UW, S3, N1, and N2 had between 1039 and 1060 unique assigned taxa at the genus level, while the number of taxa was substantially reduced at site NA (463; Table 1). At the offset of the wetland (site S1), all diverseness parameters were lower than for all other wetland sites (Table 1). Equivalent values were seen at an S1 site that was not directly associated with a wetland found (site S1R; Supplementary Tabular array S3). At site S2 in the center of the wetland, a reduction in OTU number and diversity was observed in comparison to the unpolluted wetland sites S3 and UW. The presence of Juncus plants growing within the sediment (site S2) in contrast to cottongrass (site S2C) did not pb to substantial variation in bacterial multifariousness (Supplementary Tabular array S3). Overall, the wetland maintains college taxa richness despite a pollution gradient, unlike downstream of site NA where a decrease in bacterial diversity and an increment dominance of individual taxa was observed.
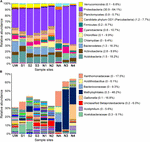
FIGURE iv. Relative abundance of bacterial taxa post-obit OTU taxonomic assignment. All assigned taxa at phylum level where possible (A) and Proteobacteria shown to the highest resolution possible down to species level where possible (B) for each sample site. Selected taxa of high abundance in multiple samples are labeled, with ranges of relative taxa affluence given in parentheses.
Proteobacteria, Actinobacteria, and Acidobacteria are commonly highly abundant in most soils, with Proteobacteria typically representing 30–40% of all bacterial taxa (Roesch et al., 2007; Lauber et al., 2009). As well, in many unlike AMD environments, these bacterial taxa dominate despite overall low diversity (Chen et al., 2016). Equally expected, Proteobacteria was dominant in all of the sites, especially in NA, N3, and N4, where this phylum accounted for 49.6%, 60.ix%, and 64.1% of the total taxon abundance, respectively (Effigy 4). Test of proteobacterial sequences in more than detail showed authority of an unclassified Xanthomonadaceae sequence particularly at site NA (17.five%) but too abundant at site S1, and an unclassified Methylophilales sequence with extremely loftier abundance (46.two%) at site N3 (Figure 4B). This unclassified Methylophilales taxon had reduced abundance at site N4 compared to N3; however, affluence of a Gallionella sp. was high (16.eight%). Members of some of these taxa have previously been associated with Fe oxidation. For example, a Xanthomonadales taxon was observed in Fe(Iii)-rich sediments (Senko et al., 2008) and inside acidophilic fe-oxidizing communities (Jones et al., 2015), although piffling is known about the organisms involved in this action. Gallionella sp. is a bacterium with well-known iron oxidizing activity in microaerophilic environments (Mitsunobu et al., 2012) capable of generating biogenic iron oxides in sediments (Kikuchi et al., 2014). Although Gallionella spp. are ordinarily reported in pH neutral environments (Gault et al., 2012), they take been previously observed at acidic mine sites (Fabisch et al., 2013; Liljeqvist et al., 2015).
The relative abundance of Proteobacteria, including Xanthomonadales, Methylophilales, and Gallionellales, was much greater in northern Afon Goch sites NA, N3, and N4 compared to other sites, while Acidobacteria, Bacteroidetes, Planctomycetes, and Verrucomicrobia were less arable in these sites (Figure 4). The ascendant taxa observed within the wetland were equivalent to those seen previously in slightly acidic, non-metallic polluted sediments of natural wetlands (Hartman et al., 2008; Peralta et al., 2013). Withal, taxa known to be involved in iron oxidation or sulfate reduction were not dominant inside the near-surface sediment wetland samples analyzed here, suggesting that they are more abundant in other locations of the AMD ecosystem, such equally deep anaerobic sediments, biofilms, or AMD mine water. For example, examination of a constructed wetland during its evolution over 225 days showed accumulation of sulfate reducing leaner and aerobic bacteria in the h2o cavalcade over time (Diaby et al., 2015). Too, samples previously taken within the southern Afon Goch wetland at the sediment–h2o interface identified taxa that grouped phylogenetically with sulfate reducing and oxidizing bacteria (Dean et al., 2013).
Factors Determining Microbial Customs Structure
Hierarchical clustering on the basis of community structure similarity found that unpolluted sites UW, N1, N2, and S3 clustered together, while a 2nd cluster included sites S1, S2, N3, and N4, with the adit site NA as an outlier (Figure 5A). There was no pregnant distinction betwixt S1 and S2 and between N3 and N4, but discrimination between wetland sites S1 and S2 compared to sites NA, N3, and N4 was pregnant (p < 0.05). On the basis of the physicochemical information, sites UW, S3, N1, and N2 were classified as an unpolluted group while sites S1, S2, N3, and N4 plus site NA were classified every bit a polluted group. ANOSIM demonstrated that there was a significant separation between the two groups (R = 0.61, p = 0.016) on the ground of community structure. NMDS ordination further demonstrated similar clustering of polluted and unpolluted sites (Figure 5B). Inclusion of the 1608 assigned taxa in the ordination indicated a greater density of taxa associated with the unpolluted sites, as shown by the taxa amassed on the left side of the plot. Of these taxa, 139 were uniquely present in the unpolluted group compared to 176 unique to the polluted group of sites. SIMPER assay indicated that the unclassified Methylophilales taxon, which was dominant at sites NA, N3, and N4 (Effigy 4B), was the highest ranked contributor to the aggregation of the sites (Effigy 5B). A Methylophilales taxon associated with Cu tolerance was previously reported as one of the nearly arable taxa within a microbial mat involved in natural remediation of heavy metal-contaminated mine h2o (Drewniak et al., 2016), while the same taxon was abundant in streams exposed to alkaline metal mine drainage (Bier et al., 2014). The other dominant taxa inside the polluted sites were as well shown to be key contributors to the blueprint of site assemblage (Figure 5B).
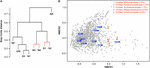
Figure v. Discrimination of sites on the basis of bacterial community structure. (A) Hierarchical clustering of sample sites based on species similarity. Sites with non-meaning clustering (p < 0.05) every bit adamant by SIMPROF are indicated with cherry-red lines. (B) Ii-dimensional NMDS plot of sites based on species similarity showing a separation betwixt unpolluted sites (UW, S3, N1, and N2), polluted sites (S1, S2, N3, and N4), and the highly polluted mine adit site (NA) on the basis of NMDS1. Assigned taxa assemblage is shown (greyness circles) and the top five taxa with the virtually contribution (% contribution) to site assemblage based on SIMPER analysis are indicated and listed in red.
Analysis of singled-out AMD environments beyond the globe has begun to identify dominant factors that influence species composition and diverseness of microbial communities exposed to AMD (Méndez-García et al., 2015; Huang et al., 2016). For many cases, pH is a ascendant factor for explaining variation in species diversity (Kuang et al., 2013; Liu et al., 2014). Indeed more generally, soil pH is oft considered as a fundamental determinant of bacterial community structure (Fierer and Jackson, 2006; Lauber et al., 2009). While nosotros observed that pH was a key cistron explaining the PCA clustering of the low pollution sites N1, N2, and S3 (Effigy 2B), BIO-ENV analysis found that pH alone was not the primal environmental variable for explaining the variation in customs construction betwixt the sites. Conductivity and metal concentration characteristics are strongly correlated with species assemblage, indicating that these factors may exist specially important (Table 2). Dissolved metal concentrations inbound the northern Afon Goch river were extremely high (upward to 628 mg L-1 of dissolved Iron at site NA) in comparing to many other studied AMD environments (Méndez-García et al., 2015), and this may in part explain the influence of electrical conductivity and dissolved Fe and other metals as a key predictor of reduced species diversity here. Analysis of an acidic Teutonic Bore mine site in Australia also observed a potent correlation between conductivity and bacterial community construction, and this location also exhibited very loftier metallic concentrations (Wakelin et al., 2012).
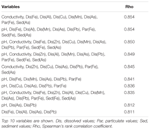
Table ii. Ready of environmental variables from BIO-ENV assay that best explain the assemblage of the bacterial community construction obtained from relative abundance of taxonomic assignments.
While site clustering on the basis of taxa assemblage could exist defined past the polluted and unpolluted groups, the wetland sites were all the same always significantly distinct from not-wetland river sites (Figure 5A). Organizing the sites as a river group and a wetland group allowed the comparison of assigned taxa on the basis of wetland establish presence. Most of the assigned taxa (73%) were shared between the river and wetland groups, while just 316 taxa were unique to the river grouping and 118 were unique to the wetland grouping. Some taxa had high abundance within wetland sites, only low abundance within not-wetland river sites (Supplementary Data Canvas 1b). These included an unclassified Bacteroidales (with an affluence of 9.8% at UW and 2.vii% at S1 but 0.one% at N3 and N4) and an unclassified Acetobacteraceae (with an affluence of eight.9% and eight.seven% at S1 and S2 but 0.three% at N1 and N2). Inside the wetland sites, there was also a large authorization of taxa incorrectly classified past the Greengenes database as "Blue-green alga," simply which include eukaryotic Stramenopiles that were identified via a chloroplast sequence, then will slightly inflate the bacterial species richness values. This is probably due to the presence of diatom species, which nosotros have previously identified from this wetland by 16S rRNA gene amplicon sequencing (Dean et al., 2013). Together, this suggests an increased dominance of photosynthetic microorganisms at the surface sediment of the wetland. In addition to plants, these oxygenic phototrophs will provide the system with oxygen and organic carbon to fuel heterotrophic activities, and will aid bulldoze biofilm product, which is an important location for sulfate metabolism to drive metal sulfide formation and metal precipitation (Roeselers et al., 2008; Chen et al., 2016).
Prediction of Metabolic Potential
Although wetland plants straight promote some bulk extraction of dissolved metals into their tissues (Dean et al., 2013), the principal bioremediation action within the wetland volition be due to microbial-derived enzymatic activities. For instance, it is clear that in that location is substantial formation of particulate metals within the middle of the wetland (Figure 2D) likely due to altered biogeochemical processes at this location (Sobolewski, 1999; Dean et al., 2013), for example, the generation of insoluble compounds during oxidation processes (Hafeznezami et al., 2012) and sulfide compounds due to metallic reduction (Wu et al., 2013), as well equally redox and pH command to farther influence metallic precipitation (Frohne et al., 2011). We might therefore expect to come across differences in functional enzymatic traits between the wetland sites and the AMD river sites. To quantify the microbial-derived metabolic potential (Kuang et al., 2016) of each site's sediment, we used the PAPRICA metabolic inference prediction method by phylogenetic placement as an culling to methodologies such equally Tax4Fun and PICRUSt, which have some limitations for environmental microbiome analysis (Bowman and Ducklow, 2015; Koo et al., 2017). Clustering of each site on the ground of metabolic potential generated a profile that distinguished NA, N3, and N4 from the other sites (Figure 6A), due to a substantial predicted decrease in abundance of many enzymes specifically at these three sites within the ochre-rich sediment (Effigy 6B). Metabolic and biochemical reactions with various elements including sulfur, arsenic, carbon, hydrogen, and nitrogen are of import for adaptation past prokaryotes in AMD habitats (Méndez-García et al., 2015). The PAPRICA model inferred decreased abundance of many of these pathways within sites NA, N3, and N4, yet some reactions, such every bit sulfate and sulfur reduction, at site NA were predicted to accept loftier abundance (Figure 6C). The overall metabolic prediction profile is particularly significant as this indicates a clear-cut stardom between the wetland and non-wetland AMD sites. The sediment at sites NA, N3, and N4 has very high Fe and As concentrations with high concentrations of other metals in the surface water. Therefore, we might speculate that the farthermost metal-rich and acidic environment leads to inhibition of some enzymatic activities within the microbial communities. The reduced metabolic potential inside these sites is not due to reduced bacterial affluence because the presented data are normalized on the basis of 16S rRNA gene amplicon sequence read number (approximately 300,000 reads taken from each site). In contrast, the metabolic potential remains stable within the wetland sediment sites despite AMD exposure. This clear departure in the metabolic potential of bacterial communities at the polluted wetland sites compared to the polluted river sites (Figure 6A) contrasts with the more subtle discrimination betwixt these sites on the ground of bacterial customs construction (Effigy 5A). We therefore propose that wetland environments with their associated AMD impacts influence the metabolic structure of sediment microbial communities. These selected communities are sufficiently tolerant of these weather that the microbial mediated processes of immobilization of metals in the sediment are sustained during the wetland evolution.
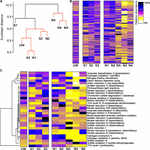
Figure half dozen. Metabolic prediction analysis and discrimination of sites on the basis of metabolic potential. (A) Hierarchical clustering of sample sites based on metabolic pathway similarity. Sites with non-significant clustering (p < 0.05) every bit determined by SIMPROF are indicated with reddish lines. Heat maps showing the predicted abundance of all prokaryotic metabolic pathways (B) and selected metabolic pathways related to element cycling (C).
An evaluation of the predicted change in affluence of enzyme classes between polluted and unpolluted sites on the southern Afon Goch (site S1 relative to site UW) and northern Afon Goch (site NA relative to site N1) was performed (Figure 7). For most enzyme classes, there was a relatively stiff positive correlation (R 2 > 0.4) in predicted enzyme abundance alter between the northern and southern river sites, such that the bulk of the enzymes that increased or decreased in abundance at site NA also increased or decreased at site S1. A 2nd analysis compared changes forth each river organisation, betwixt the site of highest source of AMD pollution (site S1 or NA) and the furthest downstream site (site S3 or N4). This provides an evaluation of metabolic potential alter moving through the wetland (x-axis) or moving down the northern Afon Goch river (y-axis; Figure 8A). These plots displayed no significant correlation (all R two < 0.1) indicating that there are very different profiles of metabolic potential depending on whether the flow is through a wetland or non. Using >two and <-two log2 fold-change as the threshold of significant enzyme abundance change, the plots predict that there are a number of significant increases and decreases in enzyme affluence through the wetland with no modify along the northern river, and as well predicted changes in enzyme abundance along the northern river that are non seen through the wetland (Figure 8A). A number of meaning increases or decreases in enzyme abundance (particularly with EC1, EC2, EC3, EC4, and EC6 enzymes) are specific to the northern river and result in no significant change through the wetland. In contrast, a few enzymes, especially of the EC1 oxidoreductase and EC2 transferase classes, have increased affluence through the wetland merely are not predicted to increase abundance between site NA and N4 (Effigy 8B). This prediction is in line with expected activities within AMD impacted wetland sediment, such as mechanisms that directly the oxidation of Fe(Two) (Sobolewski, 1999; Johnson and Hallberg, 2005; Sheoran and Sheoran, 2006), although such iron oxidation mechanisms are poorly understood and therefore challenging to predict (Ilbert and Bonnefoy, 2013). A cluster of EC3 hydrolytic enzymes are also predicted to increase abundance through the wetland (Figure 8B). Breakdown and assimilation of organic carbon products, such as cellulose and other polysaccharides from plants, volition probable require increased hydrolytic activities (Shackle et al., 2000), while metallic hydrolysis reactions will accept identify alongside oxidation reactions to remove metals from solution (Sobolewski, 1999). As metal toxicity is reduced through the wetland, metal and acidic tolerance and detoxification mechanisms are less required for survival of the microbial communities. We suggest that the observed metabolic contour indicates differential exposure of the microbiota to metal stress. Withal, it should be noted that these methods of metabolic prediction, such equally PAPRICA, Tax4Fun, and PICRUSt, may miss some functional genes, including metal tolerance genes, which are carried on plasmids.
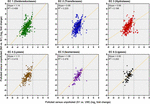
FIGURE vii. Predicted changes in enzyme abundance in response to AMD pollution. Scatter plots of Enzyme Committee (EC) enzyme reaction classes showing the relationships between wetland (southern Afon Goch) and non-wetland (northern Afon Goch) log2 fold-modify values for polluted (site S1 or NA) versus unpolluted (site UW or N1) sites. Each dot corresponds to logii fold-change in abundance of a predicted enzyme reaction. The linear regression fit line is plotted.
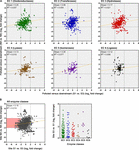
FIGURE 8. Predicted changes in enzyme abundance in response to the presence of a wetland. (A) Besprinkle plots of EC enzyme reaction classes showing the relationships betwixt wetland (southern Afon Goch) and non-wetland (northern Afon Goch) logii fold-change values for pollution source (site S1 or NA) versus downstream sites (site S3 or N4). Each dot corresponds to log2 fold-change in abundance of a predicted enzyme reaction. The linear regression fit line is plotted. (B) Scatter plot for all enzyme classes. The red-shaded quadrant indicates enzymes that are significantly increased in abundance specifically at site S3 but not at site N4, relative to the pollution source sites. The enzymes of each class that are increased in affluence at site S3 are shown (right plot).
Overview and Conclusion
Wetlands provide significant ecological functions and can provide benign ecosystem services such as promoting bioremediation of contaminated water, which may exist replicated in constructed wetlands (Zedler and Kercher, 2005; Sheoran and Sheoran, 2006; Vymazal, 2011). The potential for AMD bioremediation by wetlands derives from biogeochemical processes, which are enhanced past microbial activities (Jacob and Otte, 2003; Hafeznezami et al., 2012; Wu et al., 2013). While some of the processes controlling microbial community structure are starting time to be understood in natural wetlands with differing land apply histories, and in constructed wetlands in response to wastewater inputs (Hartman et al., 2008; Peralta et al., 2013; Ansola et al., 2014), this is the outset written report of which nosotros are aware to explore microbial customs structure dynamics within a natural wetland exposed to AMD.
We testify that a long-term AMD-impacted river, which has established a pollution-adapted wetland ecosystem, is clearly distinguished from a less adjusted, non-vegetated river on the basis of variation in microbial community structure and hence predicted office. AMD river sediment in the absence of a wetland is characterized past the prediction of significantly reduced metabolic activeness, indicative of stress response, while more specific metabolic changes are predicted within the wetland and which contribute toward the reduction in acidity and removal of dissolved metals. Our work suggests that wetlands tin can maintain high sediment microbial community diversity despite high levels of acerbity and metallic pollution, and this microbial customs structure will in turn influence biogeochemical activities that tin can take place within the wetland. Time to come investigations of these biogeochemical processes will validate the predictions of the metabolic modeling by functional analysis. Furthermore, examination of other AMD impacted natural wetlands and constructed wetlands volition be needed to place conserved functional traits inside these systems, and in particular determine whether characteristics of a long-established wetland ecosystem are also seen or tin be replicated in a recently developed constructed treatment wetland.
Information Availability
The datasets generated for this study can be plant in the European Nucleotide Archive (ENA), Study accretion number: PRJEB23187 (https://world wide web.ebi.ac.uk/ena).
Author Contributions
OA, AM, Advertizement, and JP generated the data and performed the information analysis. KW, Advert, and JP conceived the project. OA, KW, Advertizing, and JP interpreted the results and wrote the paper. All authors read and approved this manuscript.
Funding
This piece of work was financially supported in function by Ph.D. scholarship funding (to OA) from the National Council for Scientific discipline, Engineering, and Technological Innovation (CONCYTEC) of Peru and from the Faculty of Life Sciences, University of Manchester (to Advertizing and JP).
Conflict of Involvement Statement
The authors declare that the research was conducted in the absenteeism of whatsoever commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Nosotros thank Andy Hayes for MiSeq sequencing, Paul Lythgoe for ICP-AES analysis, Jennifer Nelson, Sarah Lynch, Ben Toft, and Paul Rowland for sampling work, and Matthew Cooper and Filipa Cox for communication on sequence analysis. Nosotros also thank Jon Lloyd for comments to the manuscript.
Supplementary Material
The Supplementary Cloth for this commodity tin be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01445/full#supplementary-material
References
Amaral-Zettler, L. A., Zettler, E. R., Theroux, S. Thousand., Palacios, C., Aguilera, A., and Amils, R. (2011). Microbial community structure beyond the tree of life in the extreme Rio Tinto. ISME J. 5, 42–50. doi: 10.1038/ismej.2010.101
PubMed Abstruse | CrossRef Full Text | Google Scholar
Ansola, G., Arroyo, P., and Sáenz de Miera, L. E. (2014). Characterisation of the soil bacterial community construction and composition of natural and constructed wetlands. Sci. Total Environ. 473–474(Suppl. C), 63–71. doi: 10.1016/j.scitotenv.2013.11.125
PubMed Abstract | CrossRef Total Text | Google Scholar
August, E. Eastward., McKnight, D. Yard., Hrncir, D. C., and Garhart, K. S. (2002). Seasonal variability of metals transport through a wetland impacted by mine drainage in the Rocky Mountains. Environ. Sci. Technol. 36, 3779–3786. doi: 10.1021/es015629w
CrossRef Full Text | Google Scholar
Derailed, L. C., Bakery, A. J. M., and Wheeler, B. D. (2006). The effect of vegetation on porewater composition in a natural wetland receiving acid mine drainage. Wetlands 26, 40–48. doi: 10.1672/0277-5212(2006)26[xl:TEOVOP]ii.0.CO;2
CrossRef Full Text | Google Scholar
Beining, B. A., and Otte, K. L. (1996). Retention of metals originating from an abandoned lead-zinc mine by a wetland at Glendalough, Co Wicklow. Biol. Environ. Proc. R. Ir. Acad. 96B, 117–126.
Google Scholar
Bier, R. 50., Voss, K. A., and Bernhardt, Eastward. Due south. (2014). Bacterial community responses to a gradient of element of group i mountaintop mine drainage in Central Appalachian streams. ISME J. 9, 1378–1390. doi: 10.1038/ismej.2014.222
PubMed Abstruse | CrossRef Full Text | Google Scholar
Boult, S., Collins, D. North., White, Thou. N., and Curtis, C. D. (1994). Metal send in a stream polluted by acid mine drainage - the Afon Goch, Anglesey, Uk. Environ. Pollut. 84, 279–284. doi: 10.1016/0269-7491(94)90139-ii
PubMed Abstract | CrossRef Full Text | Google Scholar
Bowman, J. S., and Ducklow, H. Due west. (2015). Microbial communities can be described by metabolic construction: a general framework and application to a seasonally variable, depth-stratified microbial community from the coastal west Antarctic Peninsula. PLoS One 10:e0135868. doi: 10.1371/periodical.pone.0135868
PubMed Abstract | CrossRef Total Text | Google Scholar
Brantner, J. Southward., and Senko, J. M. (2014). Response of soil-associated microbial communities to intrusion of coal mine-derived acid mine drainage. Environ. Sci. Technol. 48, 8556–8563. doi: ten.1021/es502261u
PubMed Abstract | CrossRef Full Text | Google Scholar
Bridge, Thousand. (2004). Contested terrain: mining and the environment. Annu. Rev. Environ. Resour. 29, 205–259. doi: x.1146/annurev.energy.28.011503.163434
CrossRef Full Text | Google Scholar
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, One thousand., Bushman, F. D., Costello, E. K., et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods seven, 335–336. doi: 10.1038/nmeth.f.303
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, L.-X., Hu, M., Huang, 50.-N., Hua, Z.-S., Kuang, J.-L., Li, S.-J., et al. (2015). Comparative metagenomic and metatranscriptomic analyses of microbial communities in acrid mine drainage. ISME J. 9, 1579–1592. doi: 10.1038/ismej.2014.245
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, L.-X., Huang, L.-N., Méndez-García, C., Kuang, J.-Fifty., Hua, Z.-Southward., Liu, J., et al. (2016). Microbial communities, processes and functions in acrid mine drainage ecosystems. Curr. Opin. Biotechnol. 38, 150–158. doi: ten.1016/j.copbio.2016.01.013
PubMed Abstruse | CrossRef Full Text | Google Scholar
Clarke, K., and Ainsworth, G. (1993). A method of linking multivariate community structure to ecology variables. Mar. Ecol. Prog. Ser. 92, 205–219. doi: 10.1016/j.biortech.2014.12.081
PubMed Abstract | CrossRef Full Text | Google Scholar
Clarke, K. R. (1993). Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18, 117–143. doi: 10.1111/j.1442-9993.1993.tb00438.x
CrossRef Full Text | Google Scholar
Clarke, K. R., Somerfield, P. J., and Gorley, R. North. (2008). Testing of null hypotheses in exploratory community analyses: similarity profiles and biota-surround linkage. J. Exp. Mar. Biol. Ecol. 366, 56–69. doi: 10.1016/j.jembe.2008.07.009
CrossRef Total Text | Google Scholar
Coupland, K., and Johnson, D. B. (2004). Geochemistry and microbiology of an impounded subterranean acidic water body at Mynydd Parys, Anglesey, Wales. Geobiology ii, 77–86. doi: ten.1111/j.1472-4677.2004.00023.x
CrossRef Full Text | Google Scholar
Dean, A. P., Lynch, S., Rowland, P., Toft, B. D., Pittman, J. One thousand., and White, K. N. (2013). Natural wetlands are efficient at providing long-term metal remediation of freshwater systems polluted by acid mine drainage. Environ. Sci. Technol. 47, 12029–12036. doi: x.1021/es4025904
PubMed Abstract | CrossRef Full Text | Google Scholar
Diaby, N., Dold, B., Rohrbach, E., Holliger, C., and Rossi, P. (2015). Temporal evolution of bacterial communities associated with the in situ wetland-based remediation of a marine shore porphyry copper tailings deposit. Sci. Total Environ. 533, 110–121. doi: 10.1016/j.scitotenv.2015.06.076
PubMed Abstract | CrossRef Full Text | Google Scholar
Drewniak, L., Krawczyk, P. S., Mielnicki, Due south., Adamska, D., Sobczak, A., Lipinski, 50., et al. (2016). Physiological and metagenomic analyses of microbial mats involved in self-purification of mine waters contaminated with heavy metals. Front. Microbiol. 7:1252. doi: 10.3389/fmicb.2016.01252
PubMed Abstruse | CrossRef Total Text | Google Scholar
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C., and Knight, R. (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200. doi: 10.1093/bioinformatics/btr381
PubMed Abstract | CrossRef Full Text | Google Scholar
Fabisch, Yard., Beulig, F., Akob, D. M., and Küsel, K. (2013). Surprising abundance of Gallionella-related iron oxidizers in creek sediments at pH 4.4 or at high heavy metallic concentrations. Front. Microbiol. 4:390. doi: 10.3389/fmicb.2013.00390
CrossRef Full Text | Google Scholar
Frohne, T., Rinklebe, J., Diaz-Bone, R. A., and Du Laing, G. (2011). Controlled variation of redox conditions in a floodplain soil: touch on on metal mobilization and biomethylation of arsenic and antimony. Geoderma 160, 414–424. doi: ten.1016/j.geoderma.2010.10.012
CrossRef Full Text | Google Scholar
Gault, A. G., Langley, Due south., Ibrahim, A., Renaud, R., Takahashi, Y., Boothman, C., et al. (2012). Seasonal changes in mineralogy, geochemistry and microbial community of bacteriogenic iron oxides (BIOS) deposited in a circumneutral wetland. Geomicrobiol. J. 29, 161–172. doi: 10.1080/01490451.2010.532196
CrossRef Full Text | Google Scholar
Hafeznezami, S., Kim, J.-Fifty., and Redman, J. (2012). Evaluating removal efficiency of heavy metals in synthetic wetlands. J. Environ. Eng. 138, 475–482. doi: 10.1061/(ASCE)EE.1943-7870.0000478
CrossRef Full Text | Google Scholar
Hallberg, Grand. B., and Johnson, D. B. (2005). Microbiology of a wetland ecosystem constructed to remediate mine drainage from a heavy metal mine. Sci. Total Environ. 338, 53–66. doi: ten.1016/j.scitotenv.2004.09.005
PubMed Abstract | CrossRef Total Text | Google Scholar
Hartman, Westward. H., Richardson, C. J., Vilgalys, R., and Bruland, G. L. (2008). Ecology and anthropogenic controls over bacterial communities in wetland soils. Proc. Natl. Acad. Sci. The statesA. 105, 17842–17847. doi: 10.1073/pnas.0808254105
PubMed Abstract | CrossRef Full Text | Google Scholar
Hedin, R. South., Narin, R. W., and Kleinmann, R. Fifty. P. (1994). Passive Treatment of Coal Mine Drainage. Washington DC: US Agency of Mines, 9389.
Google Scholar
Herlemann, D. P. R., Labrenz, Grand., Jurgens, Thousand., Bertilsson, S., Waniek, J. J., and Andersson, A. F. (2011). Transitions in bacterial communities forth the 2000 km salinity gradient of the Baltic Sea. ISME J. 5, 1571–1579. doi: 10.1038/ismej.2011.41
PubMed Abstruse | CrossRef Full Text | Google Scholar
Hua, Z.-S., Han, Y.-J., Chen, L.-X., Liu, J., Hu, M., Li, Due south.-J., et al. (2015). Ecological roles of dominant and rare prokaryotes in acrid mine drainage revealed by metagenomics and metatranscriptomics. ISME J. ix, 1280–1294. doi: 10.1038/ismej.2014.212
PubMed Abstract | CrossRef Total Text | Google Scholar
Huang, L.-Due north., Kuang, J.-Fifty., and Shu, Due west.-South. (2016). Microbial environmental and evolution in the acid mine drainage model system. Trends Microbiol. 24, 581–593. doi: x.1016/j.tim.2016.03.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Jacob, D. L., and Otte, Thou. Fifty. (2003). Conflicting processes in the wetland plant rhizosphere: metal retention or mobilization? Water Air Soil Pollut. 3, 91–104. doi: x.1023/A:1022138919019
CrossRef Full Text | Google Scholar
Johnson, D. B., and Hallberg, Thou. B. (2002). Pitfalls of passive mine water treatment. Rev. Environ. Sci. Biotechnol. 1, 335–343. doi: 10.1023/A:1023219300286
CrossRef Total Text | Google Scholar
Jones, D. South., Kohl, C., Grettenberger, C., Larson, L. N., Burgos, W. D., and Macaladya, J. L. (2015). Geochemical niches of fe-oxidizing acidophiles in acidic coal mine drainage. Appl. Environ. Microbiol. 81, 1242–1250. doi: 10.1128/AEM.02919-xiv
PubMed Abstract | CrossRef Full Text | Google Scholar
Kikuchi, Due south., Makita, H., Takai, K., Yamaguchi, N., and Takahashi, Y. (2014). Label of biogenic iron oxides collected past the newly designed liquid civilisation method using diffusion chambers. Geobiology 12, 133–145. doi: 10.1111/gbi.12073
PubMed Abstract | CrossRef Full Text | Google Scholar
Klindworth, A., Pruesse, Due east., Schweer, T., Peplies, J., Quast, C., Horn, M., et al. (2013). Evaluation of full general 16S ribosomal RNA cistron PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 41:e1. doi: ten.1093/nar/gks808
PubMed Abstract | CrossRef Full Text | Google Scholar
Koo, H., Hakim, J. A., Morrow, C. D., Eipers, P. G., Davila, A., Andersen, D. T., et al. (2017). Comparing of two bioinformatics tools used to characterize the microbial multifariousness and predictive functional attributes of microbial mats from Lake Obersee, Antarctica. J. Microbiol. Methods 140(Suppl. C), 15–22. doi: 10.1016/j.mimet.2017.06.017
PubMed Abstract | CrossRef Full Text | Google Scholar
Kuang, J., Huang, L., He, Z., Chen, Fifty., Hua, Z., Jia, P., et al. (2016). Predicting taxonomic and functional construction of microbial communities in acid mine drainage. ISME J. 10, 1527–1539. doi: 10.1038/ismej.2015.201
PubMed Abstract | CrossRef Full Text | Google Scholar
Kuang, J.-L., Huang, L.-N., Chen, Fifty.-X., Hua, Z.-S., Li, S.-J., Hu, Chiliad., et al. (2013). Contemporary environmental variation determines microbial diversity patterns in acid mine drainage. ISME J. seven, 1038–1050. doi: 10.1038/ismej.2012.139
PubMed Abstract | CrossRef Full Text | Google Scholar
Lauber, C. L., Hamady, 1000., Knight, R., and Fierer, N. (2009). Pyrosequencing-based cess of soil pH as a predictor of soil bacterial community structure at the continental calibration. Appl. Environ. Microbiol. 75, 5111–5120. doi: x.1128/AEM.00335-09
PubMed Abstract | CrossRef Full Text | Google Scholar
Liljeqvist, M., Ossandon, F. J., González, C., Rajan, South., Stell, A., Valdes, J., et al. (2015). Metagenomic analysis reveals adaptations to a common cold-adapted lifestyle in a low-temperature acid mine drainage stream. FEMS Microbiol. Ecol. 91:fiv011. doi: x.1093/femsec/fiv011
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, J., Hua, Z.-South., Chen, Fifty.-X., Kuang, J.-L., Li, Southward.-J., Shu, W.-S., et al. (2014). Correlating microbial diversity patterns with geochemistry in an extreme and heterogeneous environment of mine tailings. Appl. Environ. Microbiol. eighty, 3677–3686. doi: 10.1128/AEM.00294-14
PubMed Abstruse | CrossRef Full Text | Google Scholar
Mason, C. F. (2002). Biology of Freshwater Pollution. Essex: Longman Scientific and Technical.
Google Scholar
Mayes, W. 1000., Batty, L. C., Younger, P. 50., Jarvis, A. P., Koiv, G., Vohla, C., et al. (2009a). Wetland treatment at extremes of pH: a review. Sci. Full Environ. 407, 3944–3957. doi: 10.1016/j.scitotenv.2008.06.045
PubMed Abstract | CrossRef Full Text | Google Scholar
Mayes, Westward. M., Johnston, D., Potter, H. A. B., and Jarvis, A. P. (2009b). A national strategy for identification, prioritisation and management of pollution from abandoned not-coal mine sites in England and Wales. I. Methodology development and initial results. Sci. Total Environ. 407, 5435–5447. doi: ten.1016/j.scitotenv.2009.06.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Mayes, Westward. Yard., Potter, H. A. B., and Jarvis, A. P. (2010). Inventory of aquatic contaminant flux arising from historical metallic mining in England and Wales. Sci. Total Environ. 408, 3576–3583. doi: 10.1016/j.scitotenv.2010.04.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Méndez-García, C., Peláez, A. I., Mesa, Five., Sánchez, J., Golyshina, O. V., and Ferrer, M. (2015). Microbial diversity and metabolic networks in acid mine drainage habitats. Front. Microbiol. 6:475. doi: 10.3389/fmicb.2015.00475
CrossRef Total Text | Google Scholar
Mitsunobu, S., Shiraishi, F., Makita, H., Orcutt, B. N., Kikuchi, S., Jorgensen, B. B., et al. (2012). Bacteriogenic Atomic number 26(III) (oxyhydr)oxides characterized past synchrotron microprobe coupled with spatially resolved phylogenetic analysis. Environ. Sci. Technol. 46, 3304–3311. doi: 10.1021/es203860m
PubMed Abstruse | CrossRef Full Text | Google Scholar
Peralta, R. M., Ahn, C., and Gillevet, P. 1000. (2013). Characterization of soil bacterial customs structure and physicochemical properties in created and natural wetlands. Sci. Total Environ. 443(Suppl. C), 725–732. doi: 10.1016/j.scitotenv.2012.11.052
PubMed Abstruse | CrossRef Full Text | Google Scholar
Roesch, L. F. Westward., Fulthorpe, R. R., Riva, A., Casella, G., Hadwin, A. M. 1000., Kent, A. D., et al. (2007). Pyrosequencing enumerates and contrasts soil microbial diverseness. ISME J. 1, 283–290. doi: 10.1038/ismej.2007.53
PubMed Abstract | CrossRef Full Text | Google Scholar
Senko, J. Thousand., Wanjugi, P., Lucas, M., Bruns, Yard. A., and Burgos, W. D. (2008). Characterization of Atomic number 26(Ii) oxidizing bacterial activities and communities at ii acidic Appalachian coalmine drainage-impacted sites. ISME J. 2, 1134–1145. doi: x.1038/ismej.2008.60
PubMed Abstract | CrossRef Full Text | Google Scholar
Shackle, V. J., Freeman, C., and Reynolds, B. (2000). Carbon supply and the regulation of enzyme action in constructed wetlands. Soil Biol. Biochem. 32, 1935–1940. doi: 10.1016/S0038-0717(00)00169-3
CrossRef Full Text | Google Scholar
Sheoran, A. S., and Sheoran, V. (2006). Heavy metal removal machinery of acid mine drainage in wetlands: a disquisitional review. Miner. Eng. nineteen, 105–116. doi: 10.1016/j.mineng.2005.08.006
CrossRef Total Text | Google Scholar
Sobolewski, A. (1999). A review of processes responsible for metal removal in wetlands treating contaminated mine drainage. Int. J. Phytoremediation 1, xix–51. doi: 10.1080/15226519908500003
CrossRef Full Text | Google Scholar
Tripole, S., Gonzalez, P., Vallania, A., Garbagnati, M., and Mallea, K. (2006). Evaluation of the impact of acrid mine drainage on the chemistry and the macrobenthos in the Carolina Stream (San Luis-Argentina). Environ. Monit. Assess. 114, 377–389. doi: 10.1007/s10661-006-4941-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Wakelin, S. A., Anand, R. R., Reith, F., Gregg, A. L., Noble, R. R. P., Goldfarb, G. C., et al. (2012). Bacterial communities associated with a mineral weathering profile at a sulphidic mine tailings dump in arid Western Australia. FEMS Microbiol. Ecol. 79, 298–311. doi: ten.1111/j.1574-6941.2011.01215.ten
PubMed Abstract | CrossRef Full Text | Google Scholar
Webb, J. S., McGinness, S., and Lappin-Scott, H. G. (1998). Metal removal by sulphate-reducing bacteria from natural and constructed wetlands. J. Appl. Microbiol. 84, 240–248. doi: 10.1046/j.1365-2672.1998.00337.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Werner, J. J., Koren, O., Hugenholtz, P., DeSantis, T. Z., Walters, W. A., Caporaso, J. G., et al. (2011). Impact of training sets on classification of high-throughput bacterial 16s rRNA factor surveys. ISME J. 6, 94–103. doi: 10.1038/ismej.2011.82
PubMed Abstract | CrossRef Total Text | Google Scholar
Wu, South., Kuschk, P., Wiessner, A., Müller, J., Saad, R. A. B., and Dong, R. (2013). Sulphur transformations in synthetic wetlands for wastewater handling: a review. Ecol. Eng. 52, 278–289. doi: 10.1016/j.ecoleng.2012.eleven.003
CrossRef Full Text | Google Scholar
Zedler, J. B., and Kercher, S. (2005). Wetland resource: status, trends, ecosystem services, and restorability. Annu. Rev. Environ. Resour. thirty, 39–74. doi: 10.1146/annurev.free energy.30.050504.144248
CrossRef Total Text | Google Scholar

Do Wetlands Filter Acid Mine Pollutants,
Source: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01445/full
Posted by: wolfgodis1942.blogspot.com
0 Response to "Do Wetlands Filter Acid Mine Pollutants"
Post a Comment